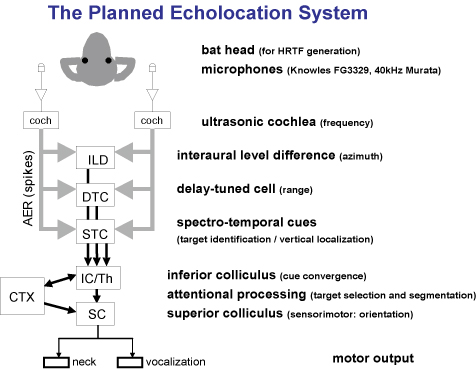
The Microchipoptera Project
Motivation:
The echolocating bat is a wonderful animal that is
unique in so many different ways: it is the only truly flying
mammal, it can use air-coupled sonar to navigate in complete darkness
while flying, it can allow its body temperature to drop to as low as
1degC (torpor) to save energy, and it has adapted to a huge range of
climates and food sources (e.g., fruits, crawling and flying insects,
nectar, fish, blood, etc). That's nice, but why would a
neuroscientist or engineer specifically study bats?
From a computational neuroscience perspective,
bats are remarkable because of the very short timescale on which they
operate. The barrage of returning sonar
echoes from a bat's near-environment lasts approximately 30 milliseconds
following a sonar emission with the echo from a specific target lasting, at
most, a few milliseconds. At this timescale, a particular neuron has the
opportunity to fire only one or two spikes to represent the echo. Unlike the “traditional” view of cortical
processing where many spikes are integrated over time to compute an average
rate, the bat must rely on populations of neurons that respond transiently but
selectively to different objects in the environment. In these neural circuits, the details of
spike timing, synaptic dynamics, and neuron biophysics become extremely
important. Flying at speeds anywhere
from 1m/s to 6 m/s, a bat’s sensory world jumps from pulse to pulse as it flies
through the world. Sensory prediction is
therefore likely to be very important in this animal. In spite of all this behavioral
specialization, the bat brain is organized like most other mammalian brains suggesting
that echolocation arises from only small modifications of the typical mammalian
auditory system.
From an engineering standpoint, biosonar systems (e.g. bats and dolphins) have
inspired the design of very sophisticated sonar and radar systems that can map
distant surfaces and track targets with great precision. Even with powerful mathematical tools and
decades of experience, however, our best systems still do not rival the
perceptual capabilities of dolphins.
Many bats demonstrate incredible aerial agility, flying in complete
darkness through branches and caves while hunting evasive insects. These animals perform such tasks in real-time
with a total power consumption (including flight) measured in Watts,
not hundreds of Watts. In addition
to the ability to navigate in complete darkness by echolocation, both bats and
dolphins live in very social environments using echolocation in group
situations without any obvious problems with interference. All of these capabilities are highly desired
by current military programs developing unmanned-aerial vehicles (UAV)
especially since many of the target environments are in places where Global
Positioning System (GPS) signals are unavailable and obstacle locations are not
mapped.
Goal:
Our goal is to construct a flying bat-sized creature
that uses ultrasonic echolocation
to both navigate and scrutinize its environment sufficiently to
distinguish
between obstacles and "insects". The bat's sensory and
motor
system will be constructed from neural models and implemented using
"neuromorphic"
VLSI techniques. Our intention is two-fold: 1) to test
these neural algorithms in a real-time, closed-loop behavioral
situation, and 2) to develop useful sonar sensors for use in miniature aircraft systems.
The Bat Head
We are working with two different hardware systems: a physically-larger
single-frequency sonar system ("narrowband") and a tiny broadband system.
The narrowband system is being used to rapidly test concepts following
initial software tests. Photos of these two systems are found below:
In the photo to the left, is our narrowband
sonar system that operates only on a frequency of 40 kHz. The fixed
arrangement of the microphones was chosen to produce a difference in echo
amplitude with azimuthal direction. The current system roughly extracts
direction and range and is capable of servoing the head (which is mounted
on an model airplane servo) to track moving targets in real-time.
On
the right, we have a photo of our broadband system using a baked polymer
clay bat head with a tiny Knowles (FG3329) microphone soldered to
the end of a group of wires. This system has two broadband ultrasonic
(and audio) microphones that will feed our silicon cochleae chips.
Both of these
physical heads produce intensity difference cues at each microphone
that allows the system to determine the angle of the arriving echo.
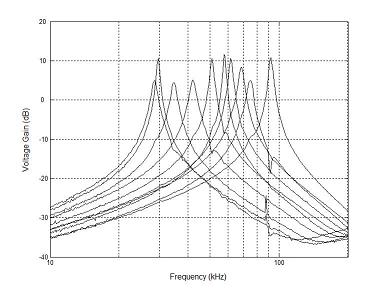
An Ultrasonic Cochlea
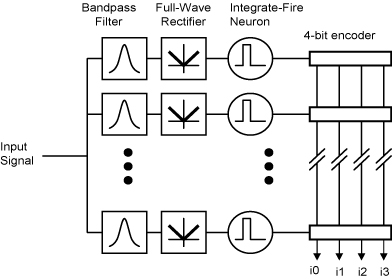
Echolocating bats specialize in high-frequency
hearing using echolocation sounds that typically range in frequency from 20 kHz
to 100 kHz. While some bats are specialized for specific
frequencies with cochlear filtering at extremely high Q10dB values, we are studying bats that use a broadband vocalization and are ultrasonic
frequency generalists (e.g., Myotis
lucifugus) with Q10dB values
in the range of 10 to 30. Good frequency resolution is important
for vertical localization, discriminating close objects, as well as for
prey discrimination.
To support our ongoing work in modeling bat echolocation,
a binaural, ultrasonic cochlea-like filter bank has been designed with moderate
quality (Q) factor (as high as 65) with spiking neurons that are driven by the filter
outputs. The neuron addresses are
reported off chip at the time of the spike in an unarbitrated fashion and in
current-mode to reduce the amount of capacitively-coupled feedback into the
filters. This chip was fabricated in a
commercially-available 0.5 um CMOS process and consumes 0.425 milliwatts
at 5 volts.
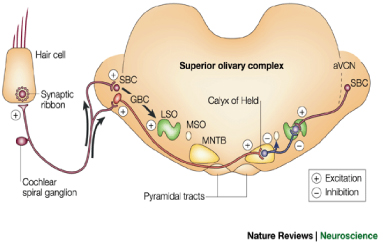
Modeling the lateral superior olive (LSO), dorsal nucleus of the lateral lemniscus (DNLL), and the inferior colliculus (IC)
When echoes arrive from different directions, the number of spikes
generated in the auditory nerve and the cochlear nucleus varies with
the intensity at each ear. Using this information, the first
binaural nucleus in the mammalian auditory system, the lateral superior
olive (or LSO) becomes selective to the direction of arrival. These cells are excited by the intensity from one ear and inhibited by the intensity from the other ear.
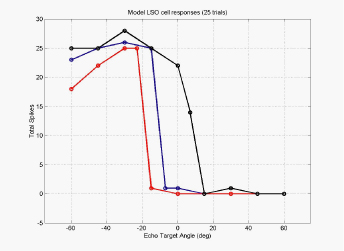
The binaural LSO response and the monaural response from the cochlear
nucleus is projected to the inferior colliculus (IC) via the doral
nucleus of the lateral lemniscus (or DNLL), resulting in very similar
responses in both DNLL and IC. With similar responses in the LSO
as in the IC, one can ask the question, "what kind of computation is
going on here?" In the figure to the left is a set of tuning
curves for three LSO cells
that have different synaptic weightings from the left and right
ears.
By comparing the responses of the population of LSO cells, each of
which have different synaptic weightings, we can determine which
direction an echo is arriving from.
In this work, we are working with a single frequency system (40kHz) and
brief burst of ultrasonic energy. Here we have designed a VLSI
circuit
that mimics the spiking neurons of the LSO, DNLL, and IC to create the
responses seen in the bat brainstem neurons.Our experimentation with
the circuits and model seem to suggest that the extra synaptic
interaction at the level of the DNLL and the IC creates an temporal
interaction between individual echoes resulting from a single
vocalization and even interactions between echoes from previous
vocalizations.
We have built a multi-layer neuron chip that can simulate the LSO, DNLL, and the IC using spiking neurons. With
this chip, we are exploring the role of long-lasting inhibition and
refractory periods on how the spatial receptive fields of bat brainstem
neurons change when multiple echoes are presented to the animal.
This is a largely unexplored area of research and where there have been
experiments, they show results that suggest that bats may need a more
sophisticated signal processing system to untangle all of the
interactions.
(see some movies in 'Multimedia')
Delay-tuned Cells (Range tuning)
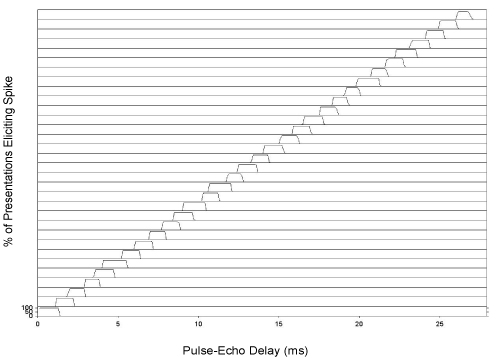
Information
about target range has many uses for bats during both prey-capture and
navigation tasks. Beyond the extraction
of distance and velocity, it may be important for less obvious tasks, such as
optimizing the parameters of the echolocation process. For example, as a bat approaches a target, it
alters the repetition rate, duration, spectral content, and amplitude of its
vocalizations. Not only is echolocation used for insect capture, it
provides to the bat information about obstacles, roosts, altitude, and other
flying creatures.
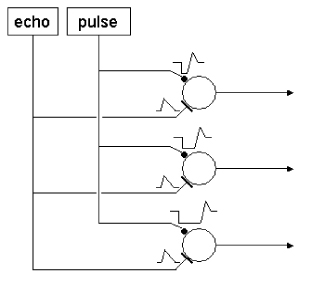
 In the bat’s
brainstem and midbrain exist neural circuits that are sensitive to the specific
difference in time between the outgoing sonar vocalization and the returning
echo. While some of the details of the
neural mechanisms are known to be species-specific, a basic model of
re-afference-triggered, post-inhibitory rebound timing is reasonably well
supported by available data.
In the bat’s
brainstem and midbrain exist neural circuits that are sensitive to the specific
difference in time between the outgoing sonar vocalization and the returning
echo. While some of the details of the
neural mechanisms are known to be species-specific, a basic model of
re-afference-triggered, post-inhibitory rebound timing is reasonably well
supported by available data.
Neurons have been
found in bats that show a ‘facilitated’ response to paired sounds (a simulated
vocalization and an echo) presented at particular delays. The cells’ responses to sounds presented at
the appropriate delays are much greater than the sum of the responses to the
individual sounds presented alone. These
cells are part of a larger class of neurons called ‘combination-sensitive’
neurons, and are specifically referred to as delay-tuned cells. Delay-tuned
cells are found at manylevels in
the bat auditory system. They have been found in the inferior
colliculus (IC), the medial geniculate body (MGB), and the auditory
cortex.
Disruption of cortical delay-tuned cells has been shown to impair a
bat’s
ability to discriminate artificial pulse-echo pair delays. It is likely
that delay-tuned neurons play a role in forming the bat’s perception of
range,
although delay-tuned cells have also been shown to respond to the
social calls
of other bats.
We have designed low-power, analog VLSI circuits
to mimic this mechanism and have demonstrated range-dependent outputs for use
in a real-time sonar system. These circuits are being used to implement
range-dependent: vocalization amplitude, vocalization rate, and closest target
isolation. Power consumption is approximately 2-4 mW.
(see some movies in 'Multimedia')
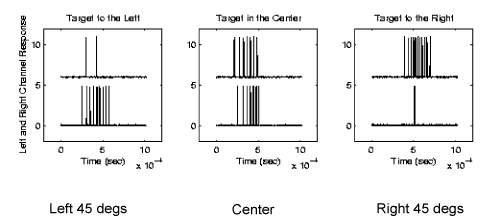
A Convergence of Range and Azimuth ( 2-D cells )
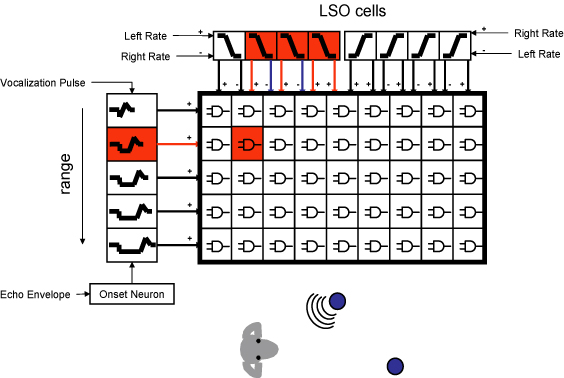
In our latest project, we have combined the work on LSO modeling which
extracts information about azimuth and combined it with the range
tuning cells above to create "2-D cells" which respond to a combination
of range and azimuth. These circuits are coincidence detectors
that respond to echoes originating from a particular direction and
range.